A história da utilização dos minerais resulta da observação dos achados arqueológicos. O homem pré-histórico, para cobrir as suas necessidades, fez uso do sílex e outras variedades de quartzo. Nas sociedades neolíticas, o homem usou gemas ( minerais utilizados em joalharia e ourivesaria ) como moeda de troca. Quando descobriu os metais ( ouro, cobre, estanho, ferro ) passou a fazer uso deles. O conhecimento dos metais e a sua utilização caracterizou alguns períodos da antiguidade, como a Idade do bronze ou a Idade do ferro. Actualmente, o homem faz uso directo ou indirecto de quase todos os minerais conhecidos, mais de 2.600 espécies minerais.
As características fundamentais de espécie mineral são a ordem geométrica, a periodicidade no arranjo da matéria, bem como a natureza dos átomos que entram na composição química da espécie mineral.
quinta-feira, 26 de março de 2009
Minerais
bacia hidrográfica
A formação da bacia hidrográfica dá-se através dos desníveis dos terrenos que orientam os cursos da água, sempre das áreas mais altas para as mais baixas.
Essa área é limitada por um divisor de águas que a separa das bacias adjacentes e que pode ser determinado nas cartas topográficas. As águas superficiais, originárias de qualquer ponto da área delimitada pelo divisor, saem da bacia passando pela seção definida e a água que precipita fora da área da bacia não contribui para o escoamento na seção considerada. Assim, o conceito de Bacia Hidrográfica pode ser entendido através de dois aspectos: rede hidrográfica e relevo.

quarta-feira, 17 de dezembro de 2008
Jgos didáticos
http://www.sobiologia.com.br/jogos/popupJogo.php?jogo=CelulaAnimal
http://www.sobiologia.com.br/jogos/popupJogo.php?jogo=flor
http://www.sobiologia.com.br/jogos/popupJogo.php?jogo=JogoDosAlimentos
Lamarckismo

Lei do Uso e do desuso
Importância do ambiente:Os seres vivos apresentam modificações que dependem do ambiente em que esses seres se desenvolvem. O ambiente condiciona a evolução, levando ao aparecimento de características que permitem aos indivíduos adapatrem-se às condições em que vivem.
De acordo com esta lei, a necessidade de um orgão num determinado ambiente, cria esse orgão e a função modifica-o, isto é, se um orgão é muito utilizado, desenvolve-se, tornando-se mais vigoroso. Se, pelo contrario, esse orgão não se usa, degenera e atrofia. Devido à dificuldade na movimentação através da vegetação densa, os órgãos locumotores das serpentes atrofiaram, resultado de uma falta de uso.
Exemplo:
Eis uma ave terrestre que é obrigada a viver em regiões inundadas ou transformadas em lagos. Levada a procurar o alimento nas águas, quer dizer, obrigada a nadar,faz esforços para este fim; po isso, afasta os dedos e a pele que une a base destes, que adquire o hábito de se distender. À força de esforços repetidos durante gerações, esta pele desenvolver-se-ia lentamente, cresceria pouco a pouco, milímetro a milímetro. Tal seria a origem da membrana interdigital, característica das patas dos gansos, dos patos e dos cisnes.

Lei da herança dos caracteres adquiridos:
Segundo esta lei, as alterações sofridas no organismo, ao longo da vida de um determinado ser, eram transmitidas aos seus descendentes por herança hereditária. Sabemos que somente por modificações nos genes é que se recebe uma herança de um antecessor, pois o DNA passa o gene para o RNA e este transfere para a proteína.
Quando o gene é transferido para a proteína não há possibilidade de modificar as informações do RNA e do DNA, portanto não existem condições para que tais alterações sejam hereditárias.
Origem da multicelularidade
Face á impossibilidade e aumentarem indefinidamente o seu volume, os seres vivos unicelulares seguiram outras vias evolutivas que haveriam de originar a organização em colónias e posteriormente a pluricelularidade. A associação de seres unicelulares em estruturas maiores e nais complexas - colónias – bem como o aparecimento de seres pluricelulares vieram permitir o aumento, mantendo elevada a superfície de contacto das células com o seu exterior. Desta forma, garantida a eficácia das trocas e a eficiência do seu metabolismo, o processo evolutivo pôde prosseguir no sentido do aumento de volume do organismo.
A alga volvox é um ser colonial constituído por uma esfera oca de células biflageladas mergulhadas numa matriz gelatinosa que as une. Os flagelos estão localizados para o exterior da esfera e permitem o movimento da colónia.
 As células mantêm a sua independência, apesar de existirem ligações citoplasmáticas entre elas. Algumas células têm função reprodutiva, o que indicia uma incipiente especialização celular. Apesar deste aumento gradual de complexidade e de interligação entre as suas células os organismos coloniais não constituem seres pluricelulares, uma vez que a diferenciação celular não existe. Admite-se que os primeiros seres multicelulares tenham surgido na sequência de um aumento de complexidade e diferenciação celular nos seres coloniais. Colónias semelhantes ao volvox, poderão, por esta via, ter dado origem ás algas verdes multicelulares. O elevado grau de interligação entre as células e a diferenciação celular abriram o caminho ao aparecimento dos tecidos e dos órgãos que caracterizam a maioria dos seres multicelulares.
As células mantêm a sua independência, apesar de existirem ligações citoplasmáticas entre elas. Algumas células têm função reprodutiva, o que indicia uma incipiente especialização celular. Apesar deste aumento gradual de complexidade e de interligação entre as suas células os organismos coloniais não constituem seres pluricelulares, uma vez que a diferenciação celular não existe. Admite-se que os primeiros seres multicelulares tenham surgido na sequência de um aumento de complexidade e diferenciação celular nos seres coloniais. Colónias semelhantes ao volvox, poderão, por esta via, ter dado origem ás algas verdes multicelulares. O elevado grau de interligação entre as células e a diferenciação celular abriram o caminho ao aparecimento dos tecidos e dos órgãos que caracterizam a maioria dos seres multicelulares.
O aparecimento da multicelularidade permitiu uma série de tendências evolutivas que acabaram por conferir vantagens aos respectivos organismos como por ex:
- a diferenciação celular, com a especialização no desempenho e de determinadas funções;
- a diminuição da taxa metabólica e utilização mais eficaz da energia.
- o aparecimento de seres maiores que mantém a constante relação superfície/volume das suas células.
- uma maior diversidade de formas que conduziu a uma melhor adaptação dos diferentes meios
- uma maior autonomia em relação ao meio externo dado que os sistemas de órgãos garantem que o meio interno mantenha um maior equilíbrio face ás flutuações do meio externo.
Notícia sobre células estaminais

Os números do Centre for International Blood and Marrow
Transplant Research (CIBMTR), mostram que 57% dos
transplantes realizados em todo o mundo são feitos
utilizando as células do próprio paciente (transplante
autólogo).
Nos restantes 43% foram utilizadas células de outros
dadores. A isto chama-se transplante alogénico. E é aqui
que reside a confusão.
A verdade é que... um transplante alogénico de células
estaminais pode ser feito a quer a partir de células de um
dador que não tenha qualquer grau de parentesco com o
paciente, através de um banco de células público, quer a
partir de células de um familiar.
Eis outro facto: os dados mostram que 64% dos
transplantes alogénicos utilizam dadores RELACIONADOS
com o paciente, e não estranhos.
Assim, e de acordo com as estatísticas mencionadas
acima, cerca de 85% de todos os transplantes feitos no
mundo utilizam quer as células estaminais do próprio
paciente, quer as de um membro da família. Estes
transplantes autólogos e entre familiares podem ser feitos
com células armazenadas em bancos privados como a
Future Health.
Ref: American National Marrow Donor Programme,


Dos procariontes ao Eucariontes
Ao longo de vários milhões de anos, os seres procariontes habitaram ambientes aquáticos e foram-se diversificando, sobretudo no que se refere ao seu metabolismo. Alguns desses seres unicelulares desenvolveram um processo metabólico que conduzia à libertação de oxigénio – a fotossíntese. O surgimento do oxigénio na atmosfera teve um impacto brutal na vida dos únicos habitantes da Terra (os procariontes). Este gás, muito reactivo, estabelece ligações com diversas moléculas, destruindo-as ou modificando-as drasticamente. Desta forma muitos grupos de procariontes foram extintos, envenenados pelo oxigénio. Contudo, alguns conseguiram sobreviver em ambientes que permaneciam anaeróbios.
Entre os sobreviventes, contam-se indivíduos que desenvolveram a capacidade de resistirem ao oxigénio. Entre eles, houve um grupo, que à semelhança das actuais mitocôndrias, era capaz de aproveitar este gás para oxidar os compostos orgânicos, obtendo assim uma grande quantidade de energia.
Apesar destas capacidades, fotossíntese e respiração, a simplicidade dos organismos procariontes limitava os processos metabólicos que podiam ser realizados simultaneamente. Alguns grupos de procariontes evoluíram e aumentaram a sua complexidade, tendo, muito provavelmente, estado na origem dos organismos eucariontes.
Fundamentalmente, existem duas hipóteses que tentam explicar a origem dos seres eucariontes a partir dos procariontes:
- Hipótese Autogénica e a
- Hipótese Endossimbiótica.
Segundo a Hipótese Autogénica, os seres eucariontes são o resultado de uma evolução gradual dos seres procariontes. Numa fase inicial, as células desenvolveram sistemas endomembranares resultantes de invaginações da membrana plasmática. Algumas dessas invaginações armazenavam o DNA, formando um núcleo. Outras membranas evoluíram no sentido de produzir organelos semelhantes ao retículo endoplasmático.
Posteriormente, algumas porções do material genético abandonaram o núcleo e evoluíram sozinhas no interior de estruturas membranares. Desta forma, formaram-se organelos como as mitocôndrias e os cloroplastos.
Esta hipótese pressupõe que o material genético do núcleo e dos organelos (sobretudo das mitocôndrias e dos cloroplastos) tenha uma estrutura idêntica. Contudo, tal não se verifica. O material genético destes organelos apresenta, geralmente, uma maior semelhança com o das bactérias autónomas, do que com o material genético presente no núcleo.
Esta e outras observações levaram ao desenvolvimento de um outro modelo ou hipótese – a Hipótese Endossimbiótica.
Esta hipótese parece reunir um maior consenso entre a comunidade científica. Esta hipótese, largamente desenvolvida por Lynn Margulis da Universidade de Massachusetts, defende que os seres eucariontes terão resultado da evolução conjunta de vários organismos procariontes, os quais foram estabelecendo associações simbióticas entre si. O termo endossimbiótica resulta do facto de algumas células viverem no interior de outras, numa relação de simbiose.
Embora este modelo admita que os sistemas endomembranares e o núcleo tenham resultado de invaginações da membrana plasmática, as mitocôndrias e os cloroplastos seriam, até à cerca de 2100 M.a., organismos autónomos. Nessa altura, algumas células de maiores dimensões (células hospedeiras) terão capturado células mais pequenas, como as ancestrais das mitocôndrias e dos cloroplastos. Alguns destes ancestrais conseguiam sobreviver no interior da célula procariótica de maiores dimensões, estabelecendo-se relações de simbiose.
A íntima cooperação entre estas células conduziu ao estabelecimento de uma relação simbiótica estável e permanente. A evolução conjunta destes organismos terá levado ao surgimento das células eucarióticas constituídas por vários organelos, alguns dos quais foram, em tempos, organismos autónomos.
Assim, as primeiras relações endossimbióticas terão sido estabelecidas com os ancestrais das mitocôndrias. Os ancestrais das mitocôndrias seriam organismos que tinham desenvolvido a capacidade de produzir energia, de forma muito rentável, utilizando o oxigénio no processo de degradação de compostos orgânicos.
Por outro lado, outro grupo de procariontes, semelhante às actuais cianobactérias, tinha desenvolvido a capacidade de produzir compostos orgânicos utilizando a energia luminosa. A associação das células procarióticas de maiores dimensões com estes seres, ancestrais dos cloroplastos, conferia-lhe vantagens evidentes.
Mas nem todas as células eucarióticas possuem cloroplastos. Este facto é explicado, segundo a Hipótese Endossimbiótica, pelo estabelecimento de relações simbióticas de forma sequencial. Isto é, as primeiras relações endossimbióticas terão sido estabelecidas com os ancestrais das mitocôndrias e, só posteriormente, algumas dessas células terão estabelecido elações de simbiose com os ancestrais dos cloroplastos.

Ciclo de vida haplodiplonte
Os indivíduos haplóides formam gâmetas, sendo por isso denominados gametófitos. A fusão de um par de gâmetas dá origem a um zigoto diplóide, que se desenvolve em um indivíduo diplóide.
A meiose, neste ciclo de vida, leva à formação de esporos e é denominada meiose espórica.
0 ciclo é chamado haplodiplonte porque existem dois tipos de indivíduo adulto, que se alternam: diplóide e haplóide. Fala-se, também, em alternância de gerações ou metagênese. 0 ciclo alternante ocorre em muitos grupos de algas. A alga verde Ulva, por exemplo, apresenta esse tipo de ciclo de vida.
Ciclo de vida diplonte
Da fusão de um par de gâmetas surge o zigoto, diplóide, que se desenvolve e origina um indivíduo diplóide, que repetirá o ciclo.
Algumas algas verdes apresentam esse tipo de ciclo, além da maior parte dos animais, inclusive nossa espécie.
Ciclo de vida haplonte
0 zigoto sofre meiose logo após se formar, produzindo células haplóides. Estas originam indivíduos haplóides, fechando o ciclo. Como a divisão reducional ocorre no zigoto, ela é denominada meiose zigótica.
O ciclo é denominado haplonte porque há, quanto à ploidia, somente um tipo de organismo adulto. A alga verde Chlamydomonas, por exemplo, tem esse tipo de ciclo de vida.
Ciclos de Vida
Tendo em conta que a reprodução sexuada apresenta dois fenómenos complementares, a meiose e a fecundação, durante o ciclo de vida de um organismo existe uma alternância entre células haplóides e diplóides.
Assim, tem-se:
- alternância de fases nucleares– a fase haplóide tem n cromossomas, ocorrendo após a meiose, e a fase diplóide tem 2n cromossomas, ocorrendo após a fecundação;
- alternância de gerações– geração é a parte do ciclo de vida de um organismo que se inicia com uma célula, dando esta origem a uma estrutura, ou entidade, multicelular, a qual irá produzir uma outra célula, através de parte da estrutura multicelular. Segundo esta definição, um ciclo de vida sexuado poderá, no máximo, apresentar duas gerações:
~geração gametófita– fase haplóide do ciclo de vida, inicia-se com o esporo e termina nos gâmetas. A estrutura multicelular da geração gametófita designa-se gametófito, onde se irão diferenciar gametângios, estruturas que contêm células que produzirão gâmetas. Os gametângios femininos designam-se oogónios (unicelulares) ou arquegónios (pluricelulares), enquanto os masculinos se designam anterídeos;
~geração esporófita - fase diplóide do ciclo, inicia-se com o zigoto e termina com a célula-mãe dos esporos. A estrutura multicelular desta fase designa-se esporófito. No esporófito diferenciam-se estruturas designadas por esporângios, contendo células que se dividem por meiose e originam esporos.
-meiose pós-zigótica - quando este fenómeno ocorre no zigoto, sendo este a única entidade diplóide do ciclo;
-meiose pré-espórica - a meiose ocorre na formação dos esporos. O zigoto, por mitoses sucessivas, origina uma entidade diplóide, onde se diferenciam células especiais que, por meiose, originam esporos;
-meiose pré-gamética - ocorre durante a formação dos gâmetas, sendo estes as únicas células haplóides do ciclo.
~~ciclo haplonte - neste tipo de ciclo de vida a fase haplóide predomina, sendo a fase diplóide constituída apenas pelo zigoto. Deste modo, considera-se que não existe verdadeira alternância de gerações. A meiose ocorre imediatamente a seguir à fecundação (meiose pós-zigótica). Este tipo de ciclo de vida é característico das algas (caminhos A e C);
~~ciclo diplonte - neste tipo de ciclo a fase diplóide predomina, sendo a fase haplóide formada apenas pelos gâmetas. Também neste caso, não existe verdadeira alternância de gerações. A meiose ocorre imediatamente antes da fecundação (meiose pré-gamética). Este tipo de ciclo é característico de animais e de algumas algas (caminhos B e D);
~~ciclo haplodiplonte - neste tipo de ciclo existe nítida alternância de fases nucleares e de gerações pois a meiose e a fecundação estão separadas no tempo. A meiose designa-se pré-espórica. Este tipo de ciclo de vida, o mais complexo, é característico das plantas superiores.
Existem, igualmente, vários tipos de reprodução sexuada:
- isogâmica - gâmetas morfologicamente iguais, geralmente flagelados, que são libertados para o meio, onde ocorre a fecundação;
- anisogâmica - gâmetas morfologicamente diferentes, sendo o feminino maior, em que ambos são libertados para o meio, existindo fecundação externa;
- oogâmica - caso particular da anisogamia, ocorre quando o gâmeta feminino é tão grande que se torna imóvel, enquanto o masculino é pequeno e móvel. Neste caso a fecundação é interna, no interior do gametângio feminino.
O monoicismo pode ser suficiente quando um indivíduo produz um zigoto por autofecundação, ou insuficiente, quando é necessária a fecundação cruzada, seja por causas morfológicas ou fisiológicas (amadurecimento desencontrado dos gâmetas, por exemplo).
Reprodução Sexuada e Meiose
Como da fecundação tem de resultar um ovo diplóide, isto é, com o número normal de cromossomas da espécie (2n), é necessário que cada gâmeta seja haplóide, ou seja, tenha metade do número destes cromossomas (n). Por este facto os gâmetas são formados através de um tipo especial de divisão celular que se chama meiose.
A meiose é, então, um processo de divisão celular a partir do qual, uma célula diplóide (2n) origina quatro células haplóides (n), isto quer dizer que, as células-filhas apresentam metade do número de cromossomas da célula-mãe.
A meiose consiste em duas divisões sucessivas, designadas por divisão I e divisão II.
Na divisão I da meiose, um núcleo diplóide (2n) origina dois núcleos haplóides (n). Pelo facto de ocorrer esta redução do número de cromossomas a divisão I também se chama de divisão reducional.
Na divisão II ocorre a separação de cromatídeos obtendo-se, assim, quatro núcleos haplóides (n), cujos cromossomas são constituídos por um cromatídeo. Pelo facto de se manter o número de cromossomas durante a divisão II esta é, também, designada de divisão equacional. A figura abaixo esquematiza como varia o número de cromossomas e a quantidade de DNA durante a meiose.
 A descrição dos principais acontecimentos da meiose permite uma melhor compreensão deste fenómeno.
A descrição dos principais acontecimentos da meiose permite uma melhor compreensão deste fenómeno.
As duas divisões nucleares que ocorrem durante a meiose são precedidas de uma única replicação do DNA. Durante o período S, os cromossomas duplicam, passando a apresentar dois cromatídeos unidos pelo centrómero.
Apresento-te, em seguida, um esquema das duas divisões da meiose e, posteriormente, a sua descrição mais detalhada.

Profase I - nesta etapa os pares de cromossomas tornam-se visíveis com longos filamentos delgados. Tal como na mitose, já foram duplicados durante a interfase precedente, logo são constituídos por dois cromatídeos unidos pelo centrómero. No entanto, nesta fase, o grau de condensação é tal que parecem estruturas unas. Os cromossomas homólogos emparelham de forma muito precisa, que se inicia em vários pontos e depois progredindo como um ziper que se fecha. Cada homólogo provém de um progenitor diferente. Este emparelhamento - sinapse - é fundamental para a ocorrência de meiose, pelo que este fenómeno não pode ocorrer em células haplóides. Nesta altura os pares de homólogos designam-se bivalentes. Durante a sinapse pedaços de cromatídeos soltam-se e voltam a ligar-se, ao acaso entre os quatro cromatídeos presentes, processo designado crossing-over. Estas trocas podem ser vistas ao microscópio pela formação de figuras em forma de X designadas quiasmas. Ao longo da Profase os quiasmas e sinapses desaparecem, tal como o nucléolo;
Metafase I - nesta etapa, tal como na mitose, o fuso acromático torna-se visível e os microtúbulos ligam-se aos centrómeros dos bivalente. Estes cromossomas emparelhados deslocam-se, então, para o centro da célula formado a placa equatorial, agora com cada centrómero do par em lados opostos da placa;
Anafase I - esta etapa inicia-se com a separação dos cromossomas homólogos, que se deslocam para pólos opostos da célula;
Telofase I - nesta etapa a espiralização dos cromossomas diminui, dando-lhes uma aparência alongada. Novas membranas nucleares são sintetizadas a partir de retículo endoplasmático rugoso enquanto se para gradualmente para a interfase. Finalmente, o fuso acromático desaparece e o nucléolo reorganiza-se. Saliente-se, no entanto, que estes acontecimentos podem não ser tão distintos, passando directamente da Telofase I para a Profase II;
Profase II - no início da segunda divisão os cromatídeos continuam unidos pelo centrómero, pelo que esta divisão se parece muito com a mitose. Se a membrana nuclear tiver sido refeita na Telofase I irá desaparecer, tal como o nucléolo, e os cromossomas irão condensar novamente;
Metafase II - forma-se novamente o fuso acromático e os cromossomas alinham-se na placa equatorial;
Anafase II - os centrómeros dividem-se e afastam-se, levados pelos microtúbulos do fuso acromático, levando os cromossomas simples para cada um dos pólos;
Telofase II - reorganização da membrana nuclear e nucléolo, com relaxação dos cromossomas, formando núcleos interfásicos.
Dou-vos também um óptimo site com a explicação por imagens da meiose: http://www.johnkyrk.com/meiosis.pt.html
Reprodução Assexuada
Neste tipo de reprodução, os descendentes desenvolvem-se a partir de uma célula ou de um conjunto de células do progenitor, pelo que todos os indivíduos são geneticamente iguais.
Assim, a partir de um só indivíduo podem formar-se numerosos indivíduos geneticamente idênticos, designando-se este agregado por clone. A produção destes indivíduos designa-se por clonagem. Todos os membros de um clone são geneticamente iguais e provêm de um só progenitor.
Só excepcionalmente podem surgir diferenças, quando por acaso ocorre uma alteração genética (mutação).
A monotonia que se verifica na descendência é consequência do processo de divisão celular que está na base da reprodução assexuada — a mitose. Este processo celular permite a formação de duas células-filhas, com uma carga hereditária exactamente igual à da célula-mãe.
Nos seres unicelulares, a mitose corresponde à própria reprodução; quando a célula se divide em duas, cada célula-filha será um novo indivíduo.
Muitos dos organismos que se reproduzem assexuadamente também o podem fazer sexuadamente, sempre que as condições do meio lhes sejam desfavoráveis. Esta capacidade permite-lhes ultrapassar o risco de extinção uma vez que a reprodução sexuada conduz à variabilidade genética e, consequentemente, a uma maior capacidade para ultrapassar a adversidade do meio ambiente. Os seres vivos em que os dois tipos de reprodução alternam periodicamente possuem alternância de gerações no seu ciclo de vida.
É ainda de referir que a mitose desempenha um papel de grande importância biológica no crescimento e desenvolvimento de seres pluricelulares, bem como na renovação tecidular. Nesta última, destaca-se a regeneração de tecidos que, nalguns organismos mais simples, pode significar a reconstrução de uma parte de um organismo ou mesmo o seu todo e, noutros organismos mais complexos, se expressa na cicatrização. Deste modo, a regeneração implica a ocorrência de divisão celular, crescimento e diferenciação.
Existem vários processos de reprodução assexuada. Os mais comuns são os seguintes: bipartição, divisão múltipla, fragmentação, gemulação, partenogénese, multiplicação vegetativa e esporulação.
Reprodução
É através da reprodução que o material genético é transmitido de geração em geração, umas vezes mantendo as características, outras produzindo algumas alterações.
A perpetuação das espécies depende da sua adaptação ao meio ambiente. Quando essa adaptação é perfeita, a reprodução deverá manter e perpetuar essas características. Porém, se, por alteração do meio, as condições deixarem de ser favoráveis, a sobrevivência das espécies estará dependente da sua capacidade de adaptação ao novo ambiente.
Para ultrapassar as incertezas do meio e assegurar a produção de novas gerações, a Natureza adoptou numerosas, e por vezes fantásticas, estratégias de reprodução, que globalmente se podem agrupar em dois processos básicos: reprodução assexuada e reprodução sexuada.
segunda-feira, 15 de dezembro de 2008
Diferenciação celular
A diferenciação celular é o processo pelo qual as células de um organismo começam tornar-se diferentes na sua forma, composição e função. A partir de então, surgem no indivíduo populações de células distintas, formando estruturas, órgãos e sistemas que interagem entre si e desempenham as diversas funções necessárias à sua sobrevivência.
Todas as células possuem um número de características fundamentais em comum; a diversidade das células pode ser vista como "variações sobre um único tema" da organização da matéria viva. Estas variações ocorrem em diferentes intensidades, de acordo com o tipo celular formado, sua função e seu grau de especialização.
As células possuem um potencial de diferenciação e um destino final; células com maior potencial são, portanto, menos diferenciadas e apresentam maiores possibilidades de diferenciação ao longo do processo. Já aquelas mais diferenciadas, ou completamente diferenciadas, perderam em grande parte ou totalmente o seu potencial.
Podemos falar de diferenciação celular em dois níveis: intracelular e intercelular.
A diferenciação intracelular refere-se às progressivas mudanças que acontecem na estrutura celular.
A diferenciação intercelular refere-se ao aparecimento de vários tipos celulares numa população de células. É o processo pelo qual duas ou mais células se tornam diferentes umas das outras.
Antes que a diferenciação aconteça, ocorre a chamada determinação, que decide o destino da célula. Pode acontecer de duas maneiras: Primeiro, pela segregação citoplasmática de moléculas determinantes no momento da clivagem do ovo, separando componentes citoplasmáticos diferentes que tornarão o citoplasma das células formadas qualitativamente diferentes. Segundo, pela indução embrionária, que envolve a interação entre células ou tecidos, condicionando células próximas a se especializarem numa determinada direcção.
Estudos feitos em embriões de tunicados mostraram que à medida que o embrião se divide, diferentes blastômeros incorporam diferentes regiões do citoplasma, que se acredita, que contenham determinantes morfogenéticos que controlam o destino da célula. Este tipo de desenvolvimento embrionário é classificado como "em mosaico", pelo facto de que as células se desenvolvem, de certa forma, de maneira independente, dividindo o embrião em regiões que darão origem a tecidos e estruturas diversas.
Também em tunicados, foi feito o isolamento de blastômeros, demonstrando a auto-diferenciação dos blastômeros isolados. No entanto, algumas interações indutivas também foram observadas; alguns tecidos como o nervoso, por exemplo, sofrem uma determinação progressiva por interação célula-célula.
Os componentes citoplasmáticos responsáveis pela determinação das células precursoras das células germinativas são os mais frequentemente estudados. Mesmo nos embriões que apresentam outros factores de regulação que não só os componentes citoplasmáticos, uma determinada região do citoplasma do ovo entrará na constituição de algumas células do embrião que certamente formarão as células germinativas.
Uma vez ocorrida a determinação, o desenvolvimento embrionário continua sendo guiado por dois fatores: a) pelos diferentes componentes do citoplasma herdados do ovo, que foram segregados anteriormente. O comportamento na diferenciação das células é, por exemplo, em vários aspectos controlado pela sua posição no embrião. Em ovos de rã e salamandra, com a entrada do espermatozóide no óvulo, iniciam-se movimentos citoplasmáticos, resultando no aparecimento de uma região oposta ao ponto de inserção do espermatozoide, "crescente cinzento", com propriedades especiais, sendo as células formadas nesta área importantes para o desenvolvimento e diferenciação celular; b) pela resposta a diferentes microambientes onde a célula se encontra, ou seja, as próprias células que a circundam, como já previsto na determinação.
Teoricamente, ainda poderíamos pensar num terceiro caminho, os diferentes tipos de genes herdados pelo indivíduo. A influência dos dois primeiros fatores (em maior ou menor grau), é que determinará como duas ou mais células fazem diferente uso da mesma informação herdada. No entanto, se estudamos o processo de diferenciação isoladamente dentro de uma mesma espécie, espera-se encontrar um padrão geral para a participação dos genes, uma vez que as células contêm o mesmo conjunto de informações herdadas.
De qualquer forma sabemos que os genes contidos no material nuclear herdado dos progenitores, expressam boa parte das características apresentadas pelos indivíduos do seu fenótipo, e que, portanto, participam activamente da diferenciação celular.
Experimentos feitos com salamandras demostraram a importância da presença do núcleo na clivagem do ovo. O isolamento, por constrição, de parte do citoplasma de um ovo não clivado resulta em um retardo temporário à clivagem na metade que não contém o núcleo. Entre os estágios de oito e dezesseis blastômeros, um dos núcleos da metade clivada, se passado para o outro lado, pode seguir a clivagem, dando origem também a um embrião normal.
Devido às várias modificações sofridas pelo núcleo no desenvolvimento embrionário, relacionadas à diferenciação celular, alguns autores utilizam o termo diferenciação nuclear ou diferenciação cromossómica para descrever alguns mecanismos peculiares observados em determinadas espécies estudadas.
Outros experimentos foram feitos com embriões de rã, removendo-se o núcleo de uma célula do teto de uma blástula ou gástrula, que ainda conservam boa parte do seu potencial, e injentando-o no citoplasma de um ovo anucleado, artificialmente activo, o ovo pôde clivar-se normalmente e desenvolver-se num girino normal. O embrião já contém, no entanto, populações distintas de células; tanto que o mesmo experimento feito com núcleos de células de outras regiões do embrião não resultou no desenvolvimento de embriões normais.
Em ovos de galinha e pato, ao isolar células de blastoderme, os seus núcleos ainda são capazes de comandar novas clivagens, formando embriões completos, porém menores.
Em plantas esta capacidade é mais expressiva, onde a maioria (senão todas) são capazes de, a partir de um pequeno grupo de células retiradas de um indivíduo adulto, formar uma nova planta completa, com todos os seus tipos celulares.
Em alguns Diptera ocorre a formação de "puffs" nos cromossomas durante a ovogênese, que são espessamentos de alguns loci específicos. Por estarem activamente ligados à síntese de RNA, indirectamente estão relacionados com a síntese de um grupo específico de proteínas que presumivelmente caracterizam um específico tipo celular.
Estes estudos indicam que a diferenciação nuclear pode ocorrer em nível de loci génicos e pode envolver a activação diferencial ou inibição desses loci pelos constituintes do microambiente onde a célula se encontra em última instância.

Ciclo celular

quarta-feira, 15 de outubro de 2008
Síntese proteica
A transcrição ocorre no interior do núcleo das células e consiste na síntese de uma molécula de mRNA(RNA Mensageiro) a partir da leitura da informação contida numa molécula de DNA. Este processo inicia-se pela ligação de um complexo enzimático à molécula de DNA, o RNA - polimerase. Esta enzima desfaz a dupla hélice, destruíndo as pontes de hidrogénio que ligam as bases complementares das duas cadeias, afastando-as.O RNA - polimerase, inicia a síntese de uma molécula de mRNA de acordo com a complementaridade das bases azotadas. Se, por exemplo, na cadeia do DNA o nucleotídeo for a adenina(A), o RNA - polimerase liga o mRNA ao nucleótido uracila(U). Quando a leitura termina, a molécula mRNA separa-se da cadeia do DNA, e esta restabelece as pontes de hidrogênio e a dupla hélice é reconstituída. Mas nem todas as sequências da molécula do DNA codificam aminoácidos.
Ao RNA sintetizado sofre um processamento ou maturação antes de abandonar o núcleo. Algumas porções do RNA transcrito, vão ser removidas - íntrons -, e as porções não removidas - éxons -, ligam-se entre si, formando assim um mRNA maturado. O RNA que sofre este processo de exclusão de porções, é designado de RNA pré-mensageiro. No final do processo, o mRNA é constituído apenas pelas sequências que codificam os aminoácidos de uma proteína, podendo assim migrar para o citoplasma, onde vai ocorrer a tradução da mensagem, isto é, a síntese de proteínas.
A tradução ocorre no citoplasma, e consiste apenas da leitura que o mRNA traz do núcleo, da qual representa uma sequência de aminoácidos, que constituí a proteína. Neste processo intervêm:
- mRNA, que vem do interior do núcleo;
- Os ribossomos;
- O tRNA (transferência);
- Enzimas (responsáveis pelo controle das reações de síntese);
- E o GTP, é o que fornece energia necessária para o processo
Nas moléculas de tRNA apresentam-se cadeias de 75 a 80 ribonucleotídeos que funcionam como intérpretes da linguagem do mRNA e da linguagem das proteínas.
O processo da tradução encerra com três etapas: iniciação, alongamento e finalização
- Iniciação: A subunidade menor do ribossomo liga-se à extremidade 5' do mRNA, esta desliza ao longo da molécua do mRNA até encontrar o códon de iniciação (AUG), transportando assim o tRNA o aminoácido metionina, ligando-se assim ao códon de iniciação por complementaridade. A subunidade maior liga-se à subunidade menor do ribossomo.
- Alongamento: Um 2º tRNA transporta um aminoácido específico de acordo com o códon. Estabelece-se uma ligação peptídica entre o aminoácido recém-chegado e a metionina. O ribossomo avança três bases ao longo do mRNA no sentido 5' -> 3', repetindo-se sempre o mesmo processo. Os tRNA que já se ligaram inicialmente, vão-se desprendendo do mRNA sucessivamente.
- Finalização: O ribossomo enconta o códon de finalização (UAA, UAG ou UGA) terminando o alongamento. O último tRNA abandona o ribossomo, as subunidades do ribossomo separam-se, podendo ser recicladas e por fim, o peptídeo é libertado.
terça-feira, 14 de outubro de 2008
Código genético

RNA

segunda-feira, 29 de setembro de 2008
Replicação do DNA

A questão da replicação do material genético foi colocada mesmo antes de Watson e Crick revelarem a estrutura do DNA. Quando estes investigadores propuseram o modelo de dupla hélice para o DNA, sugeriram uma possível forma desta molécula se replicar.
Segundo eles, a complementaridade das bases do DNA permitiria que esta molécula se autoduplicasse de forma semiconservativa: cada uma das cadeias serviria de molde para uma nova cadeia e, consequentemente, cada uma das novas moléculas de DNA seria formada por uma cadeia antiga e uma nova.
Três anos após a publicação do trabalho de Watson e Crick, um outro investigador, Arthur Kornberg, demonstrou que rea possível replicar a molécula de DNA em laboratório. Para isso bastava que, além da molécula de DNA parental, esteja presente uma enzima (DNA polimerase) e os quatro tipos de nucleótidos). Mas outros investigadores da época defendiam que a molécula de DNA apresentava dimensões demasiado elevadas para que o desenrolamento da hélice ocorresse de forma eficaz. Surgiram assim mais dois modelos que, tendo por base a complementaridade das bases do DNA, tentavam explicar o mecanismo de replicação: um modelo conservativo e um modelo dispersivo.
A hipótese conservativa admitia que a molécula de DNA progenitora se mantinha íntegra, servindo apenas de molde para a formação da molécula-filha, a qual seria formada por duas novas cadeias de nucleótidos.
Por outro lado, a hipótese dispersiva admitia que cada molécula-filha seria formada por porções da molécula inicial e por regiões sintetizadas de novo, a partir dos nucleótidos presentes na célula.




Conclusão:
Os resultados obtidos por Meselson e Stahl apoiam, inequivocamente, a hipótese semiconservativa, e podem ser interpretados da seguinte forma:
- As bactérias cultivadas em 15N incorporam esse azoto nos seus nucleótidos, formando um DNA com maior densidade, que se deposita mais próximo do fundo do tubo sujeito a centrifugação.
- Quando as bactérias são transferidas para um meio de cultura com 14N, utilizam esse azoto para produzirem novas cadeias de DNA. Assim, na primeira geração, cada molécula de DNA apresenta uma cadeia de nucleótidos com 15N (que provinha da geração parental) e outra com 14N (formada com nucleótidos que incorporaram o azoto presente no meio). Desta forma as moléculas de DNA apresentam uma densidade intermédia entre DNA com 15N e DNA com 14N.
quarta-feira, 24 de setembro de 2008

Experiência de Hershey e Chase (1952)
Antes de iniciarem as suas experiências, estes investigadores consideraram que:
- Os vírus são seres simples que não apresentam metabolismo próprio, não sendo, por isso, considerados seres vivos. - Dependem de outros seres que infectam.
- Os vírus não penetram nas células (a cápsula fica no exterior);
- As proteínas da cápsula do vírus não têm fósforo (P), mas apresentam enxofre (S);
- O DNA apresenta na sua constituição fósforo (P), mas não enxofre (S).
Isolaram, então, dois lotes de bacteriófagos (vírus que utilizam bactérias para se multiplicarem), que marcaram radioactivamente, parase conseguir seguir o seu trajectório ao microcópio. Num dos lotes, marcaram só o enxofre das proteínas (35S) e no outro somente o fósforo do DNA (32P).

Os bacteriófagos fixam-se na parede da bactéria, perfuram-na e introduzem nela o seu DNA. As proteínas da cápsula ficam no exterior. O DNA do vírus multiplica-se várias vezes no interior das bactérias. A bactéria "ao serviço do vírus" passa a produzir proteínas virais. Passado algum tempo, a parede da bactéria rompe e libertam-se para o exteior os vírus recém-formados. Como apenas o DNA viral penetra nas bactérias e não as proteínas, pode concluir-se que é o DNA que contém a informação genética necessária para a produção de novos vírus.
Experiência de Avery

Com esta experiência podemos concluir que o componente do extracto celular que constituí a substância química responsável pela transfomação dos pneumococos é o DNA dado que na amostra tratada com enzimas responsáveis pela degradação do DNA, não ocorria a transoformação das bactérias, logo, é este que contém a informação genética e que permite a transformação das bactéria de tipo S em tipo R.
Experiência de Griffith (1928)
Frederick Griffith trabalhava com bactérias da espécie Diplococcus pneumoniae, as quais provocavam pneumonia em mamíferos. Griffith verificou que esta bactéria apresentava duas formas: Tipo R, desprovidas de cápsula e com aspecto rugoso; Tipo S, envolvidas por uma cápsula de polissacarídeos que lhes confere um aspecto liso. Griffith procedeu, então, da seguinte forma:
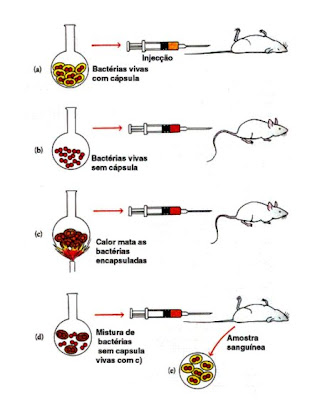
Em A injectaram-lhe bactérias do tipo S, o Rato contraiu pneumonia e morreu.
Em B injectaram-lhe bactérias do tipo R, o Rato não contraiu pneumonia e continuou saudável.
Em C injectaram-lhe bactérias do tipo S mortas, o Rato não contraiu pneumonia e morreu.
Em D injectaram-lhe uma mistura de bactérias com cápsula mortas pelo calor e bactérias vivas sem cápsula, o Rato contraiu pneumonia e morreu.
Disto conclui-se que a estipe tipo S é patogénica para os ratos, possui uma cápsula que lhe confere um aspecto liso e resistênca à fagocitose, enquanto a estripe de tipo R não é patogénica, nao possui cápsula tendo por isso um aspecto rugoso e são dstruídas pela fagocitose. Em D, o rato morre porque as estripes de tipo S transformam as características das estripes de tipo R em S, tornando ao rato impossível a sobrevivência, já que não resistem à presença da estripe S.
Ácido Desoxirribonucleico

terça-feira, 23 de setembro de 2008
Ácidos Nucleicos
Têm como função, deter e transmitir a informação genética.
Têm como unidade estrutural os nucleótidos, que unindo-se por reacção de condensação formam cadeias polinucleotídicas.
Estes são constituídos por:
- Grupo Fosfato;
- Pentose: RNA ou DNA
- Base Azotada:
- Anel simples: Citosina (C); Uracilo (exclusivo do RNA) (U); Timina (exclusivo do DNA) (T);
- Anel Duplo: Adenina (A); Guanina (G).
Prótidos
Os prótidos tem como funções: Função enzimática, estrutural, de transporte, hormonal, imunológica, motora, e de reserva alimentar.
Têm como unidade estrutural os Aminoácidos, que ligando-se etre si formam Peptidos, que ligados formam cadeias peptídicas, e estas, por fim, formam as Proteínas.
As proteínas dividem-se as seguintes estruturas:
- Estrutura primária: quando apresentam uma sequência linear de aminoácidos.
- Estrutura secundária: quando apresentam uma estrutura em hélice.
- Estrutura terciária: quando a cadeia em hélice se dobra sobre si mesma.
- Estrutura quaternária: quando as várias cadeias globulares se associam, estabelecendo interligações de hidrogénio.
Lípidos
As funções dos lípidos são: reserva energética, função estrutural, protectora vitamínica e hormonal.
Podem ser:
- Simples:
- Glicerídeos
- Triglicéridos - Complexos:
- Fosfolípidos ( Glicerol + ácido gordo + Fosfato )
Estes Lípidos simples são copostos por 1 molécula de Álcool e 3 Ácidos gordos.
Os ácidos gordos podem ser:
- Insaturados: Possuem um cadeia hidrocarbonada com uma ou mais ligações duplas.
- Saturados: Todas as ligações entre os átomos de carbbono da cadea carbonada são simples.
Glícidos
Os glícidos têm função energética e estrutural!
Glícidos podem ser:
- Monossacaídeos ou oses:
Hexoses:
- Glicose
- Frutose
- Galactose
Pentoses:
- Ribose
- Desoxirribose
- Dissacarídeos:
- Maltose = glicose + glicose
- Sacarose = glicose + frutose
- Lactose = glicose + galactose - Polissacarídeos:
- Amido (plantas)
- Glicogénio (reserva animal)
- Celulose (Fibras; paredes celulaes)
Biomoléculas
Biomoléculas podem ser:
Orgânicas(Têm carbono na sua constituição):
- Prótidos
- Lípidos
- Glícidos /glúcidos /Hidratos de carbono
- Vitaminas
- Ácidos nucleicos
Inorgânicas (não têm carbono na sua constituição):
- Água (H2O)
- Sais minerais ( Fe, K, Ca, Cl, Na, Fl)
